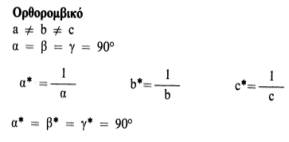Τμήμα Βιολογίας
Πανεπιστήμιο Αθηνών
Φεβρουάριος 2002
Η έννοια του κρυσταλλικού πλέγματος δίνεται στο βιβλίο «Θέματα Μοριακής
Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993. Για να εφοδιάσουμε το κρυσταλλικό
πλέγμα με την ικανότητα να περιθλά ακτίνες-Χ πρέπει να του δώσουμε κάποια
υλική υπόσταση . Ας υποθέσουμε λοιπόν ότι σε κάθε σημείο του κρυσταλλικού
πλέγματος αντιστοιχεί ένα ηλεκτρόνιο.Οι θέσεις των ηλεκτρονίων μπορούν
να καθοριστούν από τα άκρα ανυσμάτων
![]() , τέτοιων ώστε
, τέτοιων ώστε
![]() , όπου τα
, όπου τα
![]() ,
,![]() και
και
![]() είναι τα ανύσματα των αξόνων της στοιχειώδους κυψελίδας και u, v, w ακέραιοι
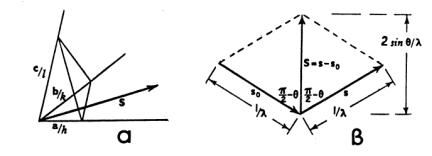
αριθμοί.Ας υποθέσουμε ότι μια παράλληλη δέσμη ακτινών-Χ μήκους κύματος
λ, "πέφτει" στο κρυσταλλικό πλέγμα κατά μια διεύθυνση που ορίζεται
από το άνυσμα
είναι τα ανύσματα των αξόνων της στοιχειώδους κυψελίδας και u, v, w ακέραιοι
αριθμοί.Ας υποθέσουμε ότι μια παράλληλη δέσμη ακτινών-Χ μήκους κύματος
λ, "πέφτει" στο κρυσταλλικό πλέγμα κατά μια διεύθυνση που ορίζεται
από το άνυσμα
![]() (βλ. Σχήμα 1).
(βλ. Σχήμα 1).
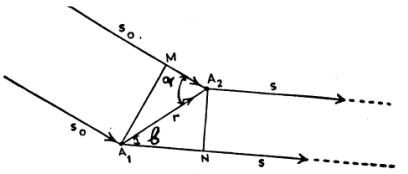
Σχήμα 1
Έστω ότι το μέτρο του ανύσματος
![]() είναι
είναι
![]() .
Τα ηλεκτρόνια εξαναγκάζονται
σε ταλάντωση και έτσι γίνονται δευτερογενείς πηγές κυμάτων.Για να βρούμε
το αποτέλεσμα συμβολής αυτών των δευτερογενών κυμάτων, είναι απαραίτητο
να πάρουμε υπ' όψη μας τις διαφορές φάσης μεταξύ των κυμάτων που σκεδάζονται
σε μια συγκεκριμένη διεύθυνση. Ας υποθέσουμε ότι επιλέγουμε μια διεύθυνση
που ορίζεται από ένα άνυσμα
.
Τα ηλεκτρόνια εξαναγκάζονται
σε ταλάντωση και έτσι γίνονται δευτερογενείς πηγές κυμάτων.Για να βρούμε
το αποτέλεσμα συμβολής αυτών των δευτερογενών κυμάτων, είναι απαραίτητο
να πάρουμε υπ' όψη μας τις διαφορές φάσης μεταξύ των κυμάτων που σκεδάζονται
σε μια συγκεκριμένη διεύθυνση. Ας υποθέσουμε ότι επιλέγουμε μια διεύθυνση
που ορίζεται από ένα άνυσμα
![]() ,που, έστω
ότι έχει επίσης μέτρο 1 / λ . Έστω δύο σημεία του κρυσταλλικού πλέγματος
Α1 και Α2 που απέχουν απόσταση
,που, έστω
ότι έχει επίσης μέτρο 1 / λ . Έστω δύο σημεία του κρυσταλλικού πλέγματος
Α1 και Α2 που απέχουν απόσταση
![]() (Υποθέτουμε ότι το Α1 επιλέγεται ως αρχή των αξόνων).Έστω Α1Μ και Α2Ν τα
μέτωπα των προσπιπτόντων και περιθλωμένων κυμάτων αντίστοιχα. Τότε, η διαφορά
δρόμων μεταξύ των δύο σκεδαζομένων κυμάτων είναι:
(Υποθέτουμε ότι το Α1 επιλέγεται ως αρχή των αξόνων).Έστω Α1Μ και Α2Ν τα
μέτωπα των προσπιπτόντων και περιθλωμένων κυμάτων αντίστοιχα. Τότε, η διαφορά
δρόμων μεταξύ των δύο σκεδαζομένων κυμάτων είναι:
![]() όπου
όπου
![]()
(αφού
![]() ,
,
![]() ,
,
![]()
και
![]() )
)
Η αντίστοιχη διαφορά φάσης είναι
![]() , αφού
, αφού
![]() .
.
Το άνυσμα
![]() λέγεται άνυσμα σκέδασης.
λέγεται άνυσμα σκέδασης.
Για να είναι τα κύματα που σκεδάζονται από τα Α1 , A2 σε φάση, θα πρέπει
η διαφορά δρόμων
![]() να ισούται με ακέραιο αριθμό μηκών
κύματος λ.Επομένως το
να ισούται με ακέραιο αριθμό μηκών
κύματος λ.Επομένως το
![]() θα πρέπει να είναι ακέραιος.
Δηλαδή,
θα πρέπει να είναι ακέραιος.
Δηλαδή,
![]() ακέραιος . Αφού αυτή η εξίσωση επαληθεύεται
όταν τα u , v , w μεταβάλλονται κατά ακέραιες τιμές, έπεται ότι τα γινόμενα
aS , bS και cSθα πρέπει να είναι ακέραιοι αριθμοί, δηλ.
ακέραιος . Αφού αυτή η εξίσωση επαληθεύεται
όταν τα u , v , w μεταβάλλονται κατά ακέραιες τιμές, έπεται ότι τα γινόμενα
aS , bS και cSθα πρέπει να είναι ακέραιοι αριθμοί, δηλ.
![]() ,
,
![]() ,
,
![]()
όπου h , k , l ακέραιοι .
Οι εξισώσεις αυτές είναι γνωστές ως εξισώσεις του Laue : Όταν
ικανοποιούνται ταυτόχρονα, δημιουργείται μια περιθλώμενη δέσμη μέγιστης
έντασης. Οι ακέραιοι h , k , l καθορίζουν την "τάξη" της περιθλώμενης
δέσμης, κατά τον ίδιο τρόπο που απλοί ακέραιοι περιγράφουν τις "τάξεις"
περίθλασης από ένα οπτικό φράγμα . Η μαθηματική μορφή των εξισώσεων Laue
(1912), αρχικά τις έκανε ακατάλληλες για ερμηνεία πειραματικών δεδομένων
μέχρις ότου ο W.L.Bragg (1913) τις τοποθέτησε σε μια φυσική βάση, με την
οποία ήταν δυνατόν να χρησιμοποιηθούν για την ερμηνεία των φασμάτων των
ακτινών-Χ και για τον καθορισμό της δομής των κρυστάλλων.
Νόμος του Bragg
Ουσιαστικά, η συνεισφορά του Bragg ήταν να συσχετίσει τους ακέραιους h , k , l των εξισώσεων Laue, με τους δείκτες Miller των κρυσταλλικών επιπέδων (βλ στο βιβλίο «Θέματα Μοριακής Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993). Μ΄ αυτό τον τρόπο έκανε αναγωγή του προβλήματος σ΄ ένα μονοδιάστατο πρόβλημα και ο τρόπος αυτός αντιμετώπισης αποδείχτηκε εξαιρετικά αποδοτικός. Η σχέση μεταξύ του νόμου του Bragg και των εξισώσεων Laue μπορεί να δειχτεί ως εξής: Οι εξισώσεις Laue μπορούν να γραφτούν:
![]() , (1)
, (1)
![]() , (2)
, (2)
![]() (3)
(3)
Αφαίρεση των (1) και (2) κατά μέλη δίνει
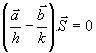 που δείχνει ότι το άνυσμα
που δείχνει ότι το άνυσμα
![]() είναι κάθετο στο άνυσμα
είναι κάθετο στο άνυσμα
![]() . Από το Σχήμα 2 είναι φανερό ότι το
. Από το Σχήμα 2 είναι φανερό ότι το
![]() είναι
ένα άνυσμα του επιπέδου με δείκτες Miller h , k , l . Με όμοιο τρόπο μπορεί
να δειχτεί ότι το άνυσμα
είναι
ένα άνυσμα του επιπέδου με δείκτες Miller h , k , l . Με όμοιο τρόπο μπορεί
να δειχτεί ότι το άνυσμα
![]() είναι κάθετο στο άνυσμα
είναι κάθετο στο άνυσμα
![]() (αφαιρώντας τις (1) και (3)) που επίσης βρίσκεται στο επίπεδο (h , k ,
l). Έτσι, το
(αφαιρώντας τις (1) και (3)) που επίσης βρίσκεται στο επίπεδο (h , k ,
l). Έτσι, το
![]() είναι κάθετο στο επίπεδο hkl . Αλλά το
είναι κάθετο στο επίπεδο hkl . Αλλά το
![]() είναι ένα άνυσμα στη διεύθυνση της διχοτόμου της
γωνίας που σχηματίζεται από την προσπίπτουσα και την περιθλώμενη δέσμη
αφού τα μέτρα των
είναι ένα άνυσμα στη διεύθυνση της διχοτόμου της
γωνίας που σχηματίζεται από την προσπίπτουσα και την περιθλώμενη δέσμη
αφού τα μέτρα των
![]() και
και
![]() είναι ίσα (βλ. Σχήμα 2). Έτσι, αυτή η διχοτόμος ταυτίζεται με την κάθετο στο επίπεδο hkl.
είναι ίσα (βλ. Σχήμα 2). Έτσι, αυτή η διχοτόμος ταυτίζεται με την κάθετο στο επίπεδο hkl.
Σχήμα 2
Αυτό είναι το πρώτο βήμα στην απόδειξη του νόμου του Bragg , και η εξήγηση
για την αντιμετώπιση κάθε περιθλώμενης δέσμης ως "ανάκλασης" από
τα επίπεδα των σημείων του κρυσταλλικού πλέγματος. Για να γίνει ο νόμος
ποσοτικός πρέπει να εισάγουμε την απόσταση dhkl των επιπέδων
hkl . Αυτή είναι η κάθετη απόσταση του επιπέδου hkl από την αρχή των αξόνων
(βλ. σχήμα) και ισούται με την προβολή του
![]() ή&
ή&
![]() ή
ή
![]() στο άνυσμα
στο άνυσμα
![]() , δηλ.
, δηλ.
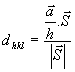
αλλά
![]()
και
![]() όπως φαίνεται εύκολα από το παραπάνω
σχήμα.
όπως φαίνεται εύκολα από το παραπάνω
σχήμα.
έτσι
![]() και
και
![]()
ή
![]()
(Νόμος του Bragg)
Η ποσότητα n που συνήθως εμφανίζεται στο νόμο του Bragg έχει "απορροφηθεί"
στους δείκτες hkl . Οι δείκτες περίθλασης σ΄ αντίθεση με τους δείκτες Miller
των επιπέδων μπορούν να έχουν κοινό παράγοντα. Εάν είναι αυτός ο κοινός
παράγοντας, η απόσταση d που αντιστοιχεί στους δείκτες nh , nk , nl
είναι το
![]() της απόστασης που αντιστοιχεί στους
δείκτες h , k , l.
της απόστασης που αντιστοιχεί στους
δείκτες h , k , l.
Αντίστροφο Πλέγμα
Η εξίσωση Bragg μπορεί να θεωρηθεί σαν αποτέλεσμα μιας φυσικής μεθόδου
λύσης των εξισώσεων Laue . Οι μαθηματικές μέθοδοι
οδηγούν σε μια διαφορετική
μορφή λύσης που εκφράζεται με την έννοια του "αντίστροφου πλέγματος"
. Αυτές οι μέθοδοι βρίσκουν τιμές του
![]() που είναι
λύσεις των εξισώσεων του Laue . Η πρώτη εξίσωση Laue δείχνει ότι η προβολή
του
που είναι
λύσεις των εξισώσεων του Laue . Η πρώτη εξίσωση Laue δείχνει ότι η προβολή
του
![]() στον
στον
![]() είναι σταθερή για ορισμένη
τιμή του h: δηλ. ότι τα άκρα των ανυσμάτων
είναι σταθερή για ορισμένη
τιμή του h: δηλ. ότι τα άκρα των ανυσμάτων
![]() , βρίσκονται
σ΄ ένα επίπεδο κάθετο στον
, βρίσκονται
σ΄ ένα επίπεδο κάθετο στον
![]() .
Εάν το h=0, το επίπεδο
περνάει από την αρχή των αξόνων, εάν το h=1 το επίπεδο τέμνει σε μια συγκεκριμένη
απόσταση τον
.
Εάν το h=0, το επίπεδο
περνάει από την αρχή των αξόνων, εάν το h=1 το επίπεδο τέμνει σε μια συγκεκριμένη
απόσταση τον
![]() ,
εάν το h=2, τον τέμνει σε διπλάσια
απόσταση, κ.ο.κ. Μ΄ άλλα λόγια ένα σύνολο επιπέδων σταθερής απόστασης δημιουργείται
και το κάθε επίπεδο του συνόλου αντιστοιχεί σε μια ορισμένη τιμή του b
.
,
εάν το h=2, τον τέμνει σε διπλάσια
απόσταση, κ.ο.κ. Μ΄ άλλα λόγια ένα σύνολο επιπέδων σταθερής απόστασης δημιουργείται
και το κάθε επίπεδο του συνόλου αντιστοιχεί σε μια ορισμένη τιμή του b
.
Με όμοιο τρόπο ένα σύνολο ισαπεχόντων επιπέδων κάθετων στον
![]() δημιουργείται και κάθε επίπεδο αντιστοιχεί σε ορισμένη τιμή του k. Ένα
τρίτο σύνολο επιπέδων αντιστοιχεί σ΄ ορισμένες τιμές του l. Οι τομείς αυτών
των επιπέδων (σημεία), αντιπροσωπεύουν τα άκρα των ανυσμάτων
δημιουργείται και κάθε επίπεδο αντιστοιχεί σε ορισμένη τιμή του k. Ένα
τρίτο σύνολο επιπέδων αντιστοιχεί σ΄ ορισμένες τιμές του l. Οι τομείς αυτών
των επιπέδων (σημεία), αντιπροσωπεύουν τα άκρα των ανυσμάτων
![]() που ικανοποιούν ταυτόχρονα τις τρεις εξισώσεις του Laue και έτσι δίνουν
τις ζητούμενες λύσεις. Το σύνολο των σημείων ορίζει ένα πλέγμα σημείων,
το "αντίστροφο πλέγμα". Τα ανύσματα του αντίστροφου πλέγματος βρίσκονται
από το γεγονός ότι καθ΄ ένα τους δημιουργείται από την τομή δύο επιπέδων
κάθετων σε δύο άξονες, έτσι το
που ικανοποιούν ταυτόχρονα τις τρεις εξισώσεις του Laue και έτσι δίνουν
τις ζητούμενες λύσεις. Το σύνολο των σημείων ορίζει ένα πλέγμα σημείων,
το "αντίστροφο πλέγμα". Τα ανύσματα του αντίστροφου πλέγματος βρίσκονται
από το γεγονός ότι καθ΄ ένα τους δημιουργείται από την τομή δύο επιπέδων
κάθετων σε δύο άξονες, έτσι το
![]() *
π.χ. είναι κάθετο στα
*
π.χ. είναι κάθετο στα
![]() και
και
![]() .
Συνεπώς, το
.
Συνεπώς, το
![]() *
μπορεί να παρασταθεί σαν p(
*
μπορεί να παρασταθεί σαν p(
![]() x
x
![]() )
όπου το p είναι μια σταθερά που πρέπει
να προσδιοριστεί. Έτσι έχουμε:
)
όπου το p είναι μια σταθερά που πρέπει
να προσδιοριστεί. Έτσι έχουμε:
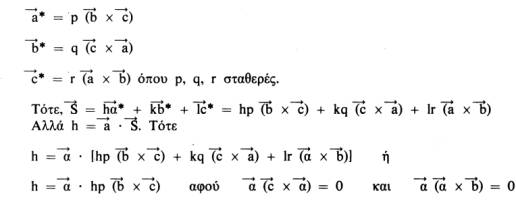
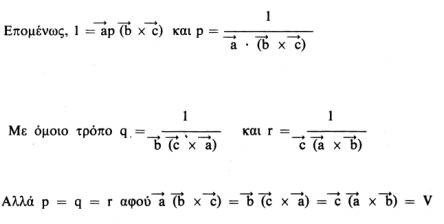

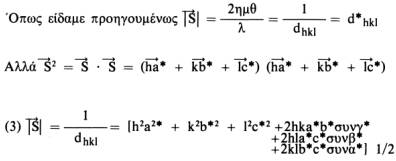
Αυτή η σχέση ισχύει για όλα τα κρυσταλλικά συστήματα και είναι ιδιαίτερα χρήσιμη. Οι σχέσεις μεταξύ των παραμέτρων του πραγματικού και του αντίστροφου πλέγματος για τα 7 κρυσταλλικά συστήματα (που διαφέρουν μεταξύ τους στο σχήμα της στοιχειώδεις κυψελίδας) είναι οι εξής:

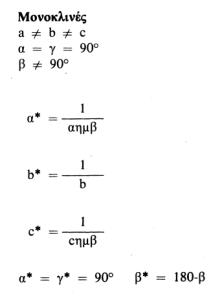 |
|
 |  |
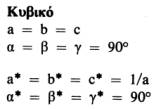 | |
|
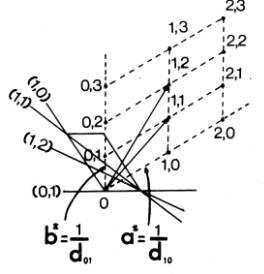
Η κατασκευή του αντίστροφου πλέγματος δείχνεται στο σχετικό απλό δισδιάστατο παράδειγμα του διπλανού σχήματος (Σχήμα 3). Έστω το πραγματικό πλέγμα (δείχνεται μόνο μια μοναδιαία κυψελίδα του) που περνάνε από τα σημεία του τα (0, 1), (1, 2), (1, 1) κ.τ.λ. Για να κατασκευάσουμε το αντίστροφο πλέγμα, παίρνουμε ένα σημείο Ο σαν αρχή των αξόνων του αντίστροφου πλέγματος (σ΄ αυτή την περίπτωση διαλέξαμε σαν σημείο Ο την αρχή των αξόνων του πραγματικού πλέγματος). |
Σχήμα 3 |
Από το σημείο αυτό σχεδιάζουμε ευθείες παράλληλες με τις καθέτους από την αρχή των αξόνων του πραγματικού πλέγματος προς τα «επίπεδα» (γραμμές) (h , k ). Παίρνουμε πάνω σ΄ αυτές τις ευθείες αποστάσεις 1/dhk, μετρούμενες από την αρχή Ο, που τα άκρα τους ορίζουν τα σημεία h ,k του αντίστροφου πλέγματος. Ενώνουμε με ευθείες (διακεκομμένες στο σχήμα) τα σημεία h k του αντίστροφου πλέγματος.
Σφαίρα Ewald ή σφαίρα ανάκλασης
Η συνθήκη για περίθλαση ακτίνων-Χ από έναν κρύσταλλο μπορεί να εκφραστεί είτε από το νόμο του Bragg, είτε από τις εξισώσεις του Laue . Ο Ewald (1921) πρότεινε τη χρήση μιας απλής γεωμετρικής κατασκευής που περιέχει και τους δύο νόμους, που απλοποιεί σημαντικά την περιγραφή των φαινομένων (Σχήμα 4).
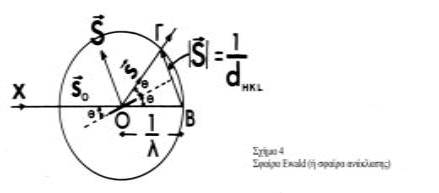
Σχεδιάζεται μια σφαίρα, ακτίνας
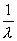 με κέντρο τον κρύσταλλο Ο. Η αρχή του αντίστροφου πλέγματος τοποθετείται στο σημείο Β όπου η προσπίπτουσα δέσμη ακτίνων-Χ, ΑΟ συναντάει την σφαίρα (σφαίρα Ewald ή σφαίρα ανάκλασης). Η συνθήκη για να παριστάνει μια ακτίνα της σφαίρας ΟΓ τη διεύθυνση των περιθλωμένων ακτίνων-Χ ικανοποιείται εφ΄ όσον το σημείο Γ είναι σημείο του αντίστροφου πλέγματος (hkl). Το άνυσμα ΒΓ σ΄ αυτή την περίπτωση είναι το άνυσμα σκέδασης
με κέντρο τον κρύσταλλο Ο. Η αρχή του αντίστροφου πλέγματος τοποθετείται στο σημείο Β όπου η προσπίπτουσα δέσμη ακτίνων-Χ, ΑΟ συναντάει την σφαίρα (σφαίρα Ewald ή σφαίρα ανάκλασης). Η συνθήκη για να παριστάνει μια ακτίνα της σφαίρας ΟΓ τη διεύθυνση των περιθλωμένων ακτίνων-Χ ικανοποιείται εφ΄ όσον το σημείο Γ είναι σημείο του αντίστροφου πλέγματος (hkl). Το άνυσμα ΒΓ σ΄ αυτή την περίπτωση είναι το άνυσμα σκέδασης
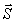 (Είναι εύκολο να δειχτεί γεωμετρικά ο νόμος του Bragg).
(Είναι εύκολο να δειχτεί γεωμετρικά ο νόμος του Bragg).
Επομένως, για να παρατηρηθεί μια, οποιαδήποτε "ανάκλαση" hkl πειραματικά, ο κρύσταλλος θα πρέπει να περιστραφεί κατά τέτοιο τρόπο ώστε το αντίστοιχο σημείο hkl του αντίστροφου πλέγματος να "πέσει" στη σφαίρα ανάκλασης.
Στα παρακάτω σχήματα δείχνεται ενδεικτικά η διαδικασία δημιουργίας του περιθλασιγράμματος ενός στρεφόμενου κρυστάλλου (η στροφή γίνεται γύρω από έναν άξονα του κρυσταλλικού πλέγματος-τα επίπεδα (στοιβάδες) του αντίστροφου πλέγματος είναι κάθετα σ΄ αυτόν) και του φιλμ που παίρνεται σε μια τέτοια διαδικασία (Σχήμα 5). Οι στοιβαδογραμμές του φιλμ δημιουργούνται όταν τα αντίστοιχα επίπεδα-στοιβάδες του αντίστροφου πλέγματος πέφτουν στη σφαίρα ανάκλασης.
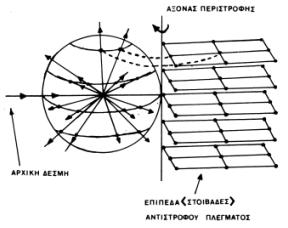 |
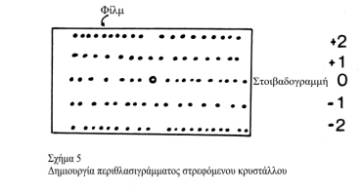 |
Νόμος του Friedel
Ο νόμος του Friedel ορίζει ότι: οι «ανακλάσεις» από τα επίπεδα hkl και
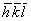 έχουν την ίδια ένταση δηλ.
έχουν την ίδια ένταση δηλ.
 ,
πράγμα που σημαίνει ότι ένα οποιοδήποτε περιθλασίγραμμα έχει κέντρο συμμετρίας (είναι κεντροσυμμετρικό). Μπορεί να δειχτεί σχετικά εύκολα, χρησιμοποιώντας τις σχέσεις που δίνονται στο βιβλίο «Θέματα Μοριακής Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993.
,
πράγμα που σημαίνει ότι ένα οποιοδήποτε περιθλασίγραμμα έχει κέντρο συμμετρίας (είναι κεντροσυμμετρικό). Μπορεί να δειχτεί σχετικά εύκολα, χρησιμοποιώντας τις σχέσεις που δίνονται στο βιβλίο «Θέματα Μοριακής Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993.
*Σημ. Ο νόμος του Friedel δεν ισχύει για ουσίες που ένα ή περισσότερα χημικά στοιχεία τους παρουσιάζουν ακμή απορρόφησης κοντά στο μήκος κύματος της χρησιμοποιούμενης ακτινοβολίας.
Περίθλαση ακτίνων-Χ από "ίνες" βιομορίων
Πολλές ουσίες μεγάλου βιολογικού ενδιαφέροντος έχουν "ινώδη" δομή στη φύση. Επιπλέον, βιομόρια μπορούν να προσανατολιστούν δημιουργώντας ίνες, όταν απομονωθούν από κύτταρα, με κατάλληλες διαδικασίες. Οι ίνες, συνήθως, αποτελούνται από επιμήκη μόρια, μόρια-αλυσίδες, πολλές φορές ελικοειδή, που πακετάρονται με τους άξονές τους παράλληλους ή σχεδόν παράλληλους με τον άξονα της ίνας. Περιθλασιγράμματα ακτίνων-Χ από τέτοιες δομές συνήθως παίρνονται τοποθετώντας τις ίνες κάθετα στη δέσμη των ακτίνων-Χ (βλ. παρακάτω σχήμα). Η διεύθυνση που είναι παράλληλη με τον άξονα της ίνας που περνάει από το κέντρο του περιθλασιγράμματος λέγεται μεσημβρινός και η κάθετη σ΄ αυτήν ισημερινός (βλ. Σχήμα 6). Εκτός από μια κοινή, μέση, αξονική διεύθυνση, η "τάξη" ή "κανονικότητα" μέσα στις ίνες μπορεί να ποικίλλει σημαντικά.
Σχήμα 6
Σε μερικές ίνες τα μόρια είναι κανονικά διευθετημένα, ώστε να δημιουργούνται κρυσταλλικές περιοχές, αλλά οι διαφορετικές κρυσταλλικές περιοχές ('κρυσταλλίτες') είναι τυχαία προσανατολισμένες γύρω από τον άξονα της ίνας (κρυσταλλικές ίνες).
Τα περιθλασιγράμματα από τέτοιες ίνες μοιάζουν με τα περιθλασιγράμματα στρεφόμενων κρυστάλλων.
(Σύγκρινε την παραπάνω φωτογραφία με το περιθλασίγραμμα στρεφόμενου κρυστάλλου).
Οι 'ανακλάσεις' βρίσκονται πάνω σε ισαπέχουσες στοιβαδογραμμές, που η απόσταση τους είναι αντίστροφα ανάλογη με την αξονική περιοδικότητα-επανάληψη της δομής: Κάθε μία στοιβαδογραμμή αντιστοιχεί σ΄ ένα επίπεδο-στοιβάδα του αντίστροφου πλέγματος. Προφανώς, περιοδικότητες κατά μήκος του άξονα της ίνας δημιουργούν 'ανακλάσεις' στον μεσημβρινό, ενώ περιοδικότητες σε προβολή κατά τον άξονα της ίνας (δηλ. σε υποθετικές εγκάρσιες τομές της ίνας) εμφανίζονται στον ισημερινό. Τα περιθλασιγράμματα όμως γίνονται πολύπλοκα, γιατί οι κηλίδες - 'ανακλάσεις' μπορούν να μετατραπούν σε τόξα (αταξία προσανατολισμού) των αξόνων των κρυσταλλιτών ως προς τον άξονα της ίνας), ή, να πλαταίνουν εφ΄ όσον οι κρυσταλλικές περιοχές είναι μικρές. Αυτό έχει σαν αποτέλεσμα οι κηλίδες να αλληλοεπικαλύπτονται, πράγμα που κάνει, πολλές φορές, ιδιαίτερα δύσκολη την ερμηνεία των περιθλασιγραμμάτων.
Πολλές ινώδεις δομές (π.χ. ινώδεις πρωτεΐνες) έχουν κανονική στερεοδιάταξη της κύριας αλυσίδας και αρκετές (π.χ. το κοινό μετάξι) δημιουργούν ψευδοκρυσταλλικές ίνες, στις οποίες η κύρια αλυσίδα επαναλαμβάνεται κανονικά, αλλά όχι και η πλευρική αλυσίδα.
Σε τέτοια περίπτωση συνηθίζεται να μιλάμε για ψευδοκυψελίδα, και όχι για πραγματική κυψελίδα, αφού μέρος μόνο της κυψελίδας επαναλαμβάνεται κανονικά σε μια τρισδιάστατη διευθέτηση. Εκτός από την ψευδοκυψελίδα, μπορεί να υπάρχει και μια αληθινή επαναλαμβανόμενη δομή, (παράλληλα με τον άξονα της ίνας (και αλυσίδας) πολύ μεγαλύτερη από την ψευδοκυψελίδα, που θα δημιουργήσει μια σειρά μεσημβρινών ανακλάσεων 'μικρής γωνίας '(low angle), κοντά δηλαδή στο κέντρο του περιθλασιγράμματος. Οι στοιβαδογράμμες αυτών των περιοδικοτήτων θα είναι πολύ κοντά η μια στην άλλη, εάν η περιοδικότητα-επανάληψη είναι μεγάλη.
Σ΄ άλλες ίνες ο βαθμός 'τάξης' είναι πολύ μικρότερος, και διάφορα είδη 'αταξιών' δημιουργούν χαρακτηριστικά φαινόμενα περίθλασης. Εάν τα μόρια είναι τυχαία μετατοπισμένα, το ένα σχετικά ως προς το άλλο, κατά τη διεύθυνση του άξονα της ίνας, κηλίδες διακριτές παρατηρούνται μόνο στον ισημερινό, ενώ οι άλλες στοιβαδογραμμές έχουν συνεχή κατανομή έντασης κατά μήκος τους (παρακρυσταλλικές ίνες). Π.χ. μια τυχαία μετατόπιση, κατά μήκος του άξονα της ίνας, κατά το μισό της επαναλαμβανόμενης περιόδου, παράγει κηλίδες διακριτές στις άρτιες στοιβαδογραμμές και συνεχή κατανομή έντασης (συνεχείς γραμμές) στης περιττής τάξης στοιβαδογραμμές.
Η αταξία ελίκωσης, που είναι συνδυασμός αταξίας περιστροφής και μετατόπισης, είναι συχνή για ελικοειδή μόρια (Μια υπεραπλούστευση της ερμηνείας παραγωγής του περιθλασιγράμματος ακτίνων-Χ ελικοειδών μορίων δίνεται στο βιβλίο «Θέματα Μοριακής Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993). Αυτού του είδους η αταξία δημιουργεί περιθλασιγράμματα με διακριτές κηλίδες στην κεντρική περιοχή και συνεχείς πλατειές κηλίδες στις στοιβαδογραμμές. Για να πάρουμε πληροφορίες για τις εντάσεις των κηλίδων που αντιστοιχούν σε ανώτερης τάξης στοιβάδες-επίπεδα του αντίστροφου πλέγματος, είναι αναγκαίο να κλίνουμε ως προς τη δέσμη των ακτίνων-Χ το δείγμα κατά το ½ της γωνίας σκέδασης 2θ έτσι ώστε το αντίστοιχο σημείο του αντίστροφου πλέγματος να 'πέσει' στη σφαίρα 'ανάκλασης' όπως φαίνεται στο παραπάνω (γ, δ) σχήμα (Κάπως έτσι ο Perutz (1951) διαπίστωσε την ύπαρξη της ανάκλασης των 1.5A που πιστοποιούσε την αλήθεια της πρότασης του L. Pauling για την ύπαρξη α-ελίκων).
Από τα περιθλασιγράμματα 'ινών' οι σταθερές του κρυσταλλικού πλέγματος μπορούν συχνά να υπολογιστούν. Μέθοδοι 'δοκιμής και σφάλματος' μπορούν να χρησιμοποιηθούν για την επίλυση της δομής.
Προπαρασκευή δειγμάτων
Μια από τις πιο σπουδαίες αλλά και δύσκολες δουλειές, σ΄ οποιαδήποτε μελέτη 'ινών' με περίθλαση ακτίνων-Χ, είναι να 'επάγουμε ' μακρiά μόρια να διευθετηθούν παράλληλα μεταξύ τους. Σε μερικές βιολογικές ίνες τα μόρια είναι 'φυσικά' προσανατολισμένα όπως προαναφέραμε όπως π.χ. στις τρίχες, κολλαγόνο, μυϊκές ίνες κ.ά.
Σ΄ άλλες περιπτώσεις, το υλικό είναι ένα διάλυμα τυχαία προσανατολισμένων μορίων και πρέπει να βρεθούν μέθοδοι για τον προσανατολισμό τους κατά ορισμένο τρόπο. 'Μακριά' μόρια μπορούν να δημιουργήσουν μια υγροκρυσταλλική φάση και είναι σπουδαίο να κατανοήσουμε τις ιδιότητες των υγρών κρυστάλλων, όταν προσπαθούμε να δημιουργήσουμε προσανατολισμένα δείγματα. Η υγροκρυσταλλική κατάσταση της ύλης είναι μια ενδιάμεση κατάσταση μεταξύ της στερεάς και της υγρής (ισοτροπικής) φάσης. Οι υγροί κρύσταλλοι 'ρέουν' όπως το H2O αλλά έχουν ανισοτροπικές ιδιότητες όπως οι στερεοί κρύσταλλοι. Όταν η συγκέντρωση ενός διαλύματος επιμηκών μορίων αυξάνει, το διάλυμα περνάει από την ισότροπη υγρή φάση, σ΄ ένα μείγμα ισορροπίας μεταξύ μιας ισοτροπικής και μιας υγροκρυσταλλικής φάσης και τελικά μεταπίπτει σε μια καθαρά υγροκρυσταλλική φάση. Οι υγροί κρύσταλλοι λέγονται αλλιώς και μεσοφάσεις ή μεσομορφικές καταστάσεις, ή ανισότροπα υγρά, ή παρακρύσταλλοι. (Πολλές βιολογικές δομές απαντούν σε υγροκρυσταλλική κατάσταση με αποτέλεσμα αυτή η φάση της ύλης να είναι εξαιρετικής σημασίας για την ίδια τη ζωή).
Οι υγροί κρύσταλλοι έχουν υποδιαιρεθεί σε διάφορους τύπους: σμηκτικούς, νηματικούς, χοληστερικούς (μια πρόσφατη διαίρεση δίνει και έναν 4ο τύπο, τους δισκοτικούς).
Στη σμηκτική μεσόφαση τα μόρια διευθετούνται έτσι ώστε τα κέντρα βάρους τους να βρίσκονται σε ισαπέχοντα επίπεδα (βλ. Σχήμα). Οι μεγάλοι άξονες των μορίων είναι κάθετοι στα επίπεδα. Τα μόρια είναι δυνατόν να κινηθούν σε δύο διαστάσεις (έτσι ώστε τα κέντρα βάρους τους να κινούνται στο ίδιο επίπεδο) είτε να περιστραφούν γύρω από τον άξονα τους. Δεν επιτρέπεται ανάμειξη μορίων διαφορετικών επιπέδων. Στη νηματική κατάσταση τα κέντρα βάρους των μορίων διευθετούνται τυχαία και οι μεγάλοι τους άξονες είναι παράλληλοι σε μια ορισμένη διεύθυνση. Περιστροφή των μορίων γύρω από το μεγάλο άξονα τους είναι επιτρεπτή (βλ. Σχήμα).
Στους στραμμένους νηματικούς ή χοληστερικούς υγρούς κρυστάλλους, τα κέντρα βάρους των μορίων σχηματίζουν ισαπέχοντα επίπεδα και τα μόρια διευθετούνται με τους μεγάλους του άξονες παράλληλους μ΄ αυτά τα επίπεδα. Σε κάθε επίπεδο τα μόρια είναι παράλληλα μεταξύ τους και σε μια κοινή διεύθυνση, αλλά από επίπεδο σε επίπεδο η διεύθυνση των μορίων μεταβάλλεται προοδευτικά και κατά σταθερό τρόπο, ώστε να δημιουργούνται ελικοειδείς διευθετήσεις μορίων, με τους άξονες των ελίκων κάθετους στα επίπεδα (βλ. Σχήμα).
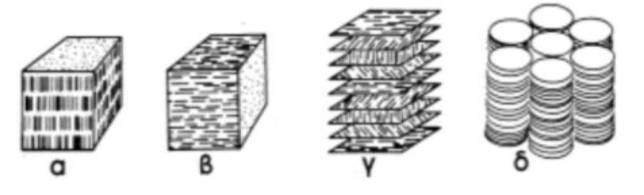
Η χοληστερική φάση βρίσκεται σε μόρια-οπτικούς αντίποδες και συνεπώς είναι σπουδαία (και εμφανίζεται συχνά ) σε μελέτες βιομορίων αφού τα βιομόρια είναι συνήθως μόρια-αντίποδες. Παράδειγμα μορίων που δημιουργούν χοληστερικές μεσοφάσεις είναι τα πολύ-α-αμινοξέα, το DNA, ο ιός της μωσαϊκής του καπνού (TMV), οι νηματοειδείς βακτηριοιοί (FV), συμπλέγματα χιτίνης-πρωτεϊνών, κυτταρίνη, διάφορες πρωτεΐνες κ.λ.π.
Στους χοληστερικούς υγρούς κρυστάλλους, η στροφή από επίπεδο σε επίπεδο μπορεί να είναι πολύ μικρή, αλλά η συνολική μακροσκοπική περιστροφή μπορεί να είναι μεγάλη. Αυτή η χοληστερική υπερδομή εμφανίζεται σαν εναλλαγή φωτεινών και σκοτεινών γραμμών, που απέχουν το μισό του βήματος της υπερέλικας, όταν το δείγμα παρατηρείται κάθετα στον άξονα της έλικας μεταξύ διασταυρωμένων, πολωτή και αναλυτή, στο πολωτικό μικροσκόπιο. Το βήμα, είναι επίσης δυνατόν, να μετρηθεί, μετρώντας τη γωνία στροφής γραμμικά πολωμένου φωτός, όταν το φως πέφτει παράλληλα με τον άξονα της έλικας. Η χοληστερική στροφή εξαρτάται από τη θερμοκρασία και από τη σχέση της διηλεκτρικής σταθεράς του διαλύτη και του μορίου, κατά μήκος του μεγάλου και των μικρών του αξόνων.
Επομένως, κατάλληλη επιλογή θερμοκρασίας ή/και διαλύτη είναι δυνατόν να ανατρέψει τη χοληστερική στροφή, επιτρέποντας στα μόρια να προσανατολίζονται καλύτερα, σε δείγματα που θα χρησιμοποιηθούν για περίθλαση ακτίνων-Χ.
Η χοληστερική συμπεριφορά μπορεί επίσης να δημιουργήσει σφαιρουλίτες, που είναι διπλοθλαστικές σφαιρικές δομές, 'βυθισμένες' στην ισότροπη (υγρή) φάση. Στους σφαιρουλίτες, ο άξονας περιστροφής των μορίων διευθύνεται κατά μια ακτίνα της σφαίρας, έτσι ώστε τα μόρια να είναι παντού εφαπτομενικά σε σφαιρικούς φλοιούς.
Ακόμα και στη νηματική φάση, παρ΄ όλο που τα μόρια, τοπικά, είναι παράλληλα το ένα με το άλλο, άλλη τοπική τάξη δεν υπάρχει, με αποτέλεσμα να υπάρχει 'αταξία' μεγάλης εμβέλειας. Επομένως, είτε τα μόρια δημιουργούν υγρούς κρυστάλλους είτε όχι, πρέπει να τα αναγκάσουμε να διευθετούνται παράλληλα το ένα με το άλλο σ΄ ολόκληρο το δείγμα, εάν θέλουμε να το χρησιμοποιήσουμε σε μελέτες περίθλασης ακτίνων-Χ.
Γι΄ αυτό το σκοπό χρησιμοποιήθηκαν πολλές μηχανικές τεχνικές:
Από πηκτώματα (gels) DNA μπορούν να 'τραβηχτούν' μακριές 'ίνες', βυθίζοντας την άκρη μιας λεπτής γυάλινης ράβδου στο πήκτωμα και αποσύροντάς την γρήγορα. Το πάχος της ίνας εξαρτάται από την ταχύτητα 'τραβήγματος'. Αργό 'τράβηγμα' μπορεί να δώσει ίνες κατάλληλες για περίθλαση ακτίνων-Χ (περίπου 0.1mm σε διάμετρο). Γρηγορότερο 'τράβηγμα' δίνει μικρότερες ίνες, που πιθανόν τα μόρια τους έχουν καλύτερη 'ευθυγράμμιση', αλλά ίσως είναι αναγκαίο να δημιουργήσουμε 'δεμάτια' απ΄ αυτές για να πάρουμε δείγματα μεγάλα, κατάλληλα για πειράματα με ακτίνες-Χ.
Διαλύματα μορίων στα οποία δεν είναι δυνατόν να εφαρμοστεί η προηγούμενη τεχνική, είναι δυνατόν να 'προσανατολιστούν', αποθέτοντας μια σταγόνα πυκνού διαλύματος μεταξύ δύο γυάλινων λεπτών ράβδων που απέχουν 1-2 mm η μια από την άλλη.
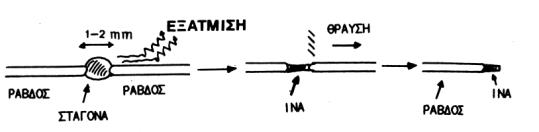
Αυτή η τεχνική χρησιμοποιείται ιδιαίτερα συχνά για πολυνουκλεοτίδια και για FV. Δείγματα πολυσακχαριτών (καθώς φυσικά και άλλων ουσιών) δημιουργήθηκαν 'αποθέτοντας' επίπεδα φυλλίδια και κατόπιν πακεττάροντας τα φυλλίδια, για τη δημιουργία ενός αρκετά 'παχέως' δείγματος. Τα δείγματα μπορεί να 'αναρτηθούν' στην άκρη μιας γυάλινης ράβδου, σε έναν κλειστό τριχοειδή σε ισορροπία με μια σταγόνα διαλύτη για σταθερή υγρασία ή μεταξύ των σταγονών μιας συσκευής που τα κρατάει υπό τάση. Μεγάλη τάση μπορεί να προκαλέσει δομικές αλλαγές στα δείγματα (π.χ. α--->β μετασχηματισμός στις κερατίνες).
Μερικά μόρια που το σχήμα τους μοιάζει με ράβδο, όπως π.χ. ο TMV, μπορούν να προσανατολιστούν 'τρέχοντας' συμπυκνωμένα διαλύματα τους μέσα από λεπτούς (0.5mm σε διάμετρο) τριχοειδείς χαλαζία.
Μερικά βιολογικά υλικά είναι διαμαγνητικά ανισότροπα και επομένως μπορούν να προσανατολιστούν σε ισχυρότατο μαγνητικό πεδίο, έτσι ώστε να δημιουργηθούν δείγματα κατάλληλα για περίθλαση ακτίνων-Χ.
Το αποτέλεσμα του μαγνητικού πεδίου σε κάθε μεμονωμένο μόριο είναι μικρό. Συνεπώς, είναι αναγκαίο να χρησιμοποιηθούν, μια υγροκρυσταλλική κατάσταση πάνω από μια κρίσιμη συγκέντρωση, όπου πολλά μόρια προσανατολίζονται σύγχρονα και πεδία μαγνητικής επαγωγής πολύ μεγάλης π.χ. πεδία που παίρνονται μ΄ ένα υπεραγώγιμο μαγνήτη (π.χ. 7 Tesla).
Τα δείγματα που έχουν προσανατολιστεί σ΄ ένα μαγνητικό πεδίο μπορούν να αποπροσανατολιστούν όταν βγουν από το πεδίο. Ο προσανατολισμός τους μπορεί να σταθεροποιηθεί 'τεντώνοντας' αργά-αργά, όταν το δείγμα βρίσκεται ακόμη στο πεδίο.
Ολόκληρα χρωμοσώματα, π.χ. τα πολυταινικά χρωμοσώματα των κυττάρων των σιελογόνων αδένων του δίπτερου εντόμου Chironomus ή της Drosophila, ή ολόκληροι ιοί, π.χ. οι ιοί FV, μπορούν να προσανατολιστούν μ΄ αυτή την τεχνική. Επίσης, ηλεκτρικά πεδία μπορούν να χρησιμοποιηθούν για τον προσανατολισμό δειγμάτων ή και συνδυασμός ηλεκτρικών και μαγνητικών πεδίων.
Σ΄ αυτή την άσκηση θα πάρετε το περιθλασίγραμμα από «φυσική» κυτταρίνη (ίνες κοινού βαμβακιού) και θα το επεξεργαστείτε. Τα τρία κύρια «σκελετικά» ή στηρικτικά συστήματα των οργανισμών περιέχουν δομικές ίνες κυτταρίνης, χιτίνης και κολλαγόνου.
Η κυτταρίνη είναι η κύρια δομική συνιστώσα των φυτών. Είτε μόνη της είτε με άλλες συνιστώσες, όπως η λιγνίνη, δημιουργεί τα κυτταρικά τοιχώματα όλων των φυτών. Το ξύλο, ουσιαστικά αποτελείται από «άδεια» κυτταρικά τοιχώματα από κυτταρίνη και λιγνίνη. Το μόριο της κυτταρίνης, ένας πολυσακχαρίτης, είναι μια μακριά, αδιακλάδωτη αλυσίδα, από 14.000 περίπου κατάλοιπα γλυκόζης (Μ.Β.~2.3x106daltons) που συνδέονται μεταξύ τους με β-1,4 γλυκοζιδικούς δεσμούς (βλ. Σχήμα)
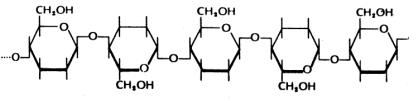
Τα μόρια της κυτταρίνης οργανώνονται και δημιουργούν ινίδια που με την σειρά τους, πακετάρονται με διάφορους τρόπους σε διαφορετικούς ιστούς και δημιουργούν, πολλές φορές, δομές εξαιρετικής αντοχής (πολλές φορές ανθεκτικότερες από το ατσάλι).
Η χιτίνη είναι η κύρια δομική συνιστώσα των μυκήτων και του εξωσκελετού (επιδερμίδιου) των αρθροπόδων (καβουριών και εντόμων). Κι' αυτή είναι ένα γραμμικό, μη διακλαδιζόμενο πολυμερές, ενός παραγώγου της γλυκόζης της Ν-ακετυλογλουκοζαμίνης (NAG). (Ο χημικός τύπος του NAG δείχνεται στο βιβλίο «Θέματα Μοριακής Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993).
Διαδοχικά κατάλοιπα NAG συνδέονται και σ' αυτή την περίπτωση με β-1,4- γλυκοζιδικούς δεσμούς. Συχνότατα η χιτίνη απαντάται σε συνδυασμό με πρωτεΐνες. Στα ζώα , η κύρια «σκελετική» δομή είναι το σύστημα του κολλαγόνου αλλά συχνά και η χιτίνη αποτελεί ένα κύριο, χαρακτηριστικό, στηρικτικό υλικό στα ασπόνδυλα. Διερωτάται κανείς γιατί τα συστήματα κυτταρίνης και χιτίνης δεν είναι κατάλληλα και δεν χρησιμοποιούνται σαν συνδετικοί ιστοί στα ζώα.
Μια πιθανή εξήγηση είναι ότι το κολλαγόνο μπορεί να τροποποιηθεί άμεσα με μια πολύ ειδική κολλαγενάση (ένζυμο), ενώ ένα σύστημα χιτίνης-πρωτεϊνών π.χ. το δερμάτιο των εντόμων (επιδερμίδιο) μπορεί να τροποποιηθεί μόνο από πρωτεάσες, που θα μπορούσαν να προσβάλλουν το προστατευτικό στρώμα των πρωτεϊνών που είναι συνδεδεμένες με την χιτίνη. Ίσως, να μη βρέθηκε τρόπος, κατά τη διάρκεια της εξέλιξης , να πετυχαίνεται κάτι τέτοιο ειδικά, χωρίς να επηρεάζονται άλλες πρωτεΐνες του συνδετικού ιστού. Το δείγμα που θα χρησιμοποιηθεί στην άσκηση αποτελείται από «ίνες» κοινού βαμβακιού (κυτταρίνης), προσανατολισμένες περίπου παράλληλα μεταξύ τους.
Η διαγραμματική παράσταση της συσκευής περίθλασης ακτίνων-Χ που θα χρησιμοποιηθεί στην άσκηση δείχνεται στο παρακάτω σχήμα:
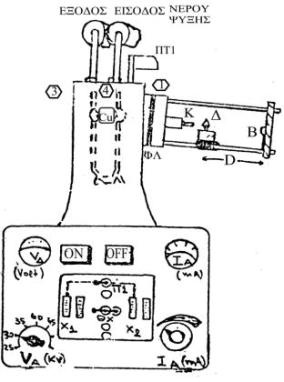
|
ON/OFF:Διακόπτης Λειτουργίας Συσκευής VΔ:Τάση δικτύου (220V) VΑ:Ρυθμιστής ανοδικής Τάσης ΙΑ:Ρυθμιστής ανοδικού Ρεύματος (ΜΑ) και μιλλιαμπερόμετρο Χ1:Χρονόμετρο Λεπτών Χ2:Χρονόμετρο Ωρών Π1:Παράθυρο 1 (ρύθμιση) Θχ:Θέση Διακόπτη Χρόνου Cu:Λυχνία Cu ΠΤ1:΄Aνοιγμα / Κλείσιμο Παραθύρου 1 ΦΛ:Φίλτρο Νi (λ= 1.542 A) Κ:Κατευθυντήρας (Collimator) Δ:Θέση (Υποδοχή) Δείγματος Β:Παγίδα (Backstop) Φ:Θέση film Ο:Απόσταση Δείγματος-film |
|
Συσκευή περίθλασης ακτίνων-Χ, που βρίσκεται στο Ευρωπαϊκό Κέντρο Μοριακής Βιολογίας (EMBL) της Χαϊδελβέργης |
 |
Οι ίνες βαμβακιού τοποθετούνται οριζόντιες και κάθετες στη δέσμη των ακτίνων-Χ στη θέση υποδοχής του δείγματος Δ, όπως φαίνεται στο παρακάτω διάγραμμα.
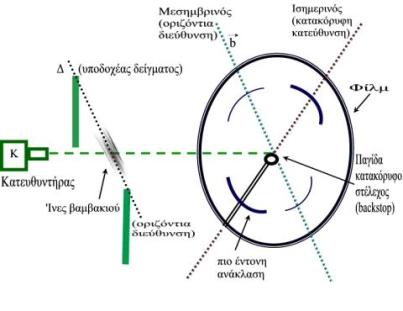
Επομένως οι διευθύνσεις του ισημερινού και μεσημβρινού του φιλμ, όπως ορίστηκαν παραπάνω, είναι αυτές που δείχνονται στο διάγραμμα για τη γεωμετρία του πειράματος.
Δίνεται ότι:
- Οι κρυσταλλίτες της κυτταρίνης του βαμβακιού είναι κρυσταλλίτες του μονοκλινούς συστήματος, με σταθερές πλέγματος (στοιχειώδους κυψελίδας).
a = 8.35A
b = 10.30A
c = 7.90A
β = 84ο
(α=γ=90ο)
- Επιπλέον, δίνεται ότι ο b άξονας είναι περίπου παράλληλος με τον άξονα των ινών.
Βοήθημα: Επομένως οι ανακλάσεις 0κ0 (δηλ. 010, 020 κ.ο.κ.) θα πρέπει να εμφανίζονται στην παραπάνω γεωμετρία, κατά μήκος του μεσημβρινού (Σκεφτείτε επομένως πού θα εμφανίζονται οι άλλες 'ανακλάσεις').
- Ζητείστε οπωσδήποτε από τον υπεύθυνο της άσκησης την απόσταση δείγματος - φιλμ. Θα σας χρειαστεί.
- Το μήκος κύματος της ακτινοβολίας που χρησιμοποιείται είναι λ=1.542 A (φιλτραρισμένη με φίλτρο Ni CuKα).
Διαδικασία λήψης του περιθλασιγράμματος
1. Αναγνωρίστε με προσοχή τα τμήματα της συσκευής.
2. Τοποθετείτε το δείγμα στη θέση υποδοχής του, όπως υποδείχτηκε προηγουμένως.
3. Γεμίστε τον υποδοχέα με φιλμ ακτίνων-Χ και τοποθετήστε τον σωστά στη συσκευή.
4. Ρυθμίστε το χρόνο έκθεσης του φιλμ (1-2 ώρες), την απόσταση δείγματος-φιλμ, και τον προσανατολισμό του δείγματος.
5. Αφού ρυθμίσετε την παροχή του νερού ψύξης της αντικαθόδου, ρυθμίστε τις συνθήκες λειτουργίας (διαφορά δυναμικού μεταξύ καθόδου-ανόδου, ανοδική τάση), (κουμπί VA), και ρεύμα ανοδικό (κουμπί ΙΑ) σε τιμές και με τρόπο που θα σας υποδειχτεί από τον υπέυθυνο. Μη ξεχάσετε (!) ν΄ ανοίξετε το παράθυρο ακτίνων-Χ, έχοντας προηγουμένως τοποθετήσει το κατάλληλο φίλτρο Νi.
Προσοχή: το μολύβδινο προστατευτικό κάλυμμα να είναι σωστά τοποθετημένο. Πάρτε τις κατάλληλες προφυλάξεις (!). Βεβαιωθείτε ότι η παγίδα (back-stop) είναι τοποθετημένη σωστά.
6. Μετά την έκθεση του φιλμ τον κατάλληλο χρόνο, εμφανίστε το στο σκοτεινό θάλαμο, στερεώστε-ξεπλύνατε-στεγνώστε το.
(Όλες αυτές οι διαδικασίες να γίνουν με μεγάλη προσοχή).
| Ενδεικτικό περιθλασίγραμμα ακτίνων-Χ που έχει προέλθει από «ίνες» κοινού βαμβακιού (κυτταρίνη) |  |
Αποτίμηση του περιθλασιγράμματος
1. Χρησιμοποιώντας τις σταθερές του κρυσταλλικού πλέγματος της κυτταρίνης που θα σας δόθηκαν (a=8.35A, b=10.3A, c=7.9A, β=84ο) και τον τύπο:
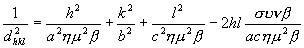
που ισχύει για το μονοκλινές σύστημα (όπως εύκολα μπορεί να δειχτεί από τις εξισώσεις που σας δόθηκαν προηγουμένως (σελ. 7-8) υπολογίστε τα
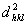 για όσες ανακλάσεις (hkl) νομίζετε αναγκαίο.
για όσες ανακλάσεις (hkl) νομίζετε αναγκαίο.
Προσοχή: Μην ξεχνάτε ότι τα h, k, l μπορούν να πάρουν και αρνητικές τιμές (π.χ. μπορείτε να έχετε την ανάκλαση
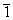 0 1).
0 1).
2. Κατασκευάστε έναν πίνακα της μορφής που δίνεται παρακάτω και συμπληρώστε τη στήλη
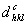 ,
με τις τιμές που υπολογίσατε.
,
με τις τιμές που υπολογίσατε.
| (A) |
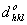 (A) (A) |
3. Φτιάξτε ενδεικτικά ένα διάγραμμα του περιθλασιγράμματος που πήρατε, σημειώνοντας προσεκτικά θέση, σχήμα και ένταση (ενδεικτικά) των 'ανακλάσεων' που πιθανόν διακρίνετε.
4. Μπορείτε να υπολογίσετε από το φιλμ, το
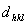 για κάθε 'ανάκλαση'-κηλίδα hkl ως εξής:
για κάθε 'ανάκλαση'-κηλίδα hkl ως εξής:
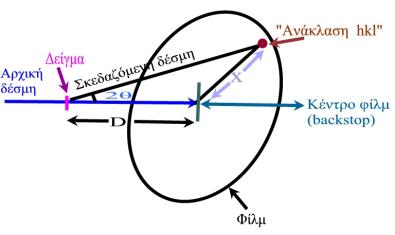
Από το σχήμα φαίνεται ότι
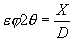 όπου D η απόσταση δείγματος-φιλμ και Χ η απόσταση 'ανάκλασης' (κηλίδας) κέντρου περιθλασιγράμματος. (Αφού σε κάθε 'ανάκλαση' hkl υπάρχει και η κεντροσυμμετρική της hkl, σύμφωνα με το νόμο του Friedel, καλύτερο είναι να μετρήσετε την απόσταση 'ανάκλασης'-κεντροσυμμετρικής της και να διαιρέσετε με το 2 για να υπολογίσετε το Χ).
όπου D η απόσταση δείγματος-φιλμ και Χ η απόσταση 'ανάκλασης' (κηλίδας) κέντρου περιθλασιγράμματος. (Αφού σε κάθε 'ανάκλαση' hkl υπάρχει και η κεντροσυμμετρική της hkl, σύμφωνα με το νόμο του Friedel, καλύτερο είναι να μετρήσετε την απόσταση 'ανάκλασης'-κεντροσυμμετρικής της και να διαιρέσετε με το 2 για να υπολογίσετε το Χ).
Από το νόμο του Bragg
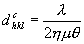 (λ~1.542A).
(λ~1.542A).
5. Παίρνοντας υπ΄ όψη τον προσανατολισμό του δείγματος, και τα
που μετρήσατε, συγκρίνοντας τα με τα
του πίνακα μπορείτε να βρείτε τους δείκτες h, k, l κάθε κυλίδας-'ανάκλασης'.
Στο ενδεικτικό διάγραμμα (3 πιο πάνω) που φτιάξατε (που θα πρέπει να είναι αρκετά ρεαλιστικό) σημειώστε πάνω σε κάθε 'ανάκλαση' τους δείκτες της h, k, l.
Συμπληρώστε στον πίνακα (βλ. 2 πιο πάνω) τις τιμές
που μετρήσατε και δίπλα την ένταση τους Ι (αμαύρωση) ενδεικτικά. (Παραστήστε με 5 αστερίσκους (*****) την ισχυρότερη και 1 αστερίσκο την ασθενέστερη και ανάλογα τις ενδιάμεσες.
Σημ. Εάν δεν ξέρατε τις σταθερές της κυψελίδας (και φυσικά το κρυσταλλικό σύστημα) θα έπρεπε να τις βρείτε έχοντας σαν μοναδικά δεδομένα τα
των ανακλάσεων (όπως και έγινε αρχικά, φυσικά).
Απαντήστε κατά τον καλύτερο δυνατό τρόπο, στις παρακάτω ερωτήσεις:
1) Από τις σχετικές εντάσεις των ανακλάσεων, μήπως μπορείτε να βγάλετε συμπεράσματα για τον -'κατά προσέγγιση'-προσανατολισμό του μορίου στη στοιχειώδη κυψελίδα: (Μην ξεχνάτε ότι οι δέκτες hkl παριστάνουν και επίπεδα).
2) Με ποιά διαδικασία θα προχωρούσατε σε κατασκευή μοντέλου της κυτταρίνης και έλεγχό του (συμβουλευτείτε το βιβλίο «Θέματα Μοριακής Βιοφυσικής», Σ. Ι. Χαμόδρακας, Αθήνα 1993).
3) Σε τι είδους 'αταξίας' νομίζετε ότι οφείλεται η πλάτυνση των ισημερινών 'ανακλάσεων'-κηλίδων του περιθλασιγράμματος.
4) Πώς νομίζετε ότι θα μπορούσατε να βελτιώσετε το περιθλασίγραμμα που πήρατε: (επεμβαίνοντας στο δείγμα).
Στους υπεύθυνους θα παραδώσετε:
a) Μια σύντομη περιγραφή όλης της διαδικασίας.
b) Το φιλμ που πήρατε.
c) Το διάγραμμα στο οποίο κάνατε δεικτοδότηση των ανακλάσεων, και τον πίνακα των υπολογισμών.
d) Απαντήσεις στις ερωτήσεις.
ΒΙΒΛΙΟΓΡΑΦΙΑ:
1. 'The Determination of Crystal Structures' (The Crystalline State, Vol. III) (1968) Lipson, H. and Cochran, W., G. Bell and Sons Ltd., London.
2. 'Diffraction of X-rays by Proteins, Nucleic Acids and Viruses' (1966). Wilson, H.R, E. Arnold, London.
3. 'X-ray Fibre Diffraction' (1982) Marvin, D.A. and Nave, C. in 'Current Methods in Structure Molecular Biology', Plenum Press.
4. 'Cell Structure and Function' (1973) Lowey, A.G. and Siekevitz, P., Holt, Rinehart and Winston, London.
5. 'The Chitin System' (1973) Biol. Rev. 49, pp 597-636 Rudal, K.M. and Kenchington, W.